Chapter Two
The Fungi of Woodlands
'He that high grouth on cedars did bestowe
Gave also lowly mushrumpes leave to growe'
-- Robert Southwell, Spirituall Poems, Grosart, 69, 1595.
The variety of situations in which fungi grow provides clues to the character of their food requirements and reveals something of the way in which these are satisfied. The superficial covering of 'mould' that appears so readily upon bread, cheese, leather, or other organic materials left about in damp situations, consists mainly of the spore-bearing branches of the mould fungi from which currents of air constantly waft away myriads of spores. These branches grow up vertically from a dense web of mycelium distributed through the bread or other material. The mildews that sometimes disfigure the leaves and other parts of plants with a cobweb-like growth in damp weather are parasitic fungi of similar habit. The hyphae composing the vegetative mycelium form and excrete substances that attack chemically the starch, sugar, or other organic constituents of the dead or living substrates, breaking them down into simpler and more soluble substances that can be absorbed by the hyphae and used as food. Similarly, the bracket or mushroom-like sporophores that appear upon living trees, dead stumps, and felled timber are the spore-bearing organs of fungi whose vegetative mycelium is parasitic upon the living tissues of the wood and other parts of the tree, or saprophytic upon those of the dead stumps or timber on which the sporophores are borne. Not infrequently, the spore-bearing organs of such parasitic fungi are produced upon the dead parts of trees still living, appearing only after the active mycelium responsible has killed the branch from which they emerge (Pl. 1, 6, 7).
The mushrooms, toadstools, and puff-balls that grow so plentifully on the ground in pastures and woodlands are the sporophores of many different species of fungi, the vegetative mycelium of each of which is living and growing on organic matter in the soil beneath the surface. In some cases, as in the truffle fungi, the sporophores not only develop below ground but remain underground when mature.
In order to understand how the mycelium of these soil fungi lives and grows and the part played in the soil by their activities as a whole, a short digression is desirable.
Every gardener knows that soil consists of a mixture of mineral particles -- sand, clay, chalk, and the like -- mixed with organic materials in various stages of decay, and that it is the well-decomposed part of the latter that gives its dark colour and crumbly texture to fertile garden soil. If he be a wise gardener, he knows also that this organic material or humus is a great reservoir of fertility for plant growth, and he is constantly adding to it by suitable manures or by plant remains from his compost heap.
In natural soils, the stock of humus is continually renewed by the death and gradual decay of the bodies or parts of bodies of animals and plants living in the soil and on its surface.
But in cultivated soils, much of the potential humus-forming material is removed with successive crops and it is one of the main objects of good husbandry to restore it in suitable forms. Fresh supplies of organic residues help to keep the soil open and well aerated, provide food for a most varied assortment of animals and plants during its conversion to humus, and eventually, in the form of the dark-coloured crumbly substance so-called, improves the texture of the soil for plant growth, increases its water-holding capacity, and constitutes a valuable reserve stock of potential food of vital importance to healthy growth and soil fertility.
In addition to the surface vegetation that it carries, it must not be forgotten that the soil is the home of a teeming population of animals and plants, ranging from relatively large mobile creatures such as earthworms and insects, to vast numbers of plants and animals of microscopic size forming collectively what is usually called the soil microflora and microfauna. Fungi and bacteria are specially well represented in the former and the activities of members of these two groups play an important part not only in bringing about the changes that lead to the formation of humus, but also in many other complicated processes by which the fertility of soil for the growth of ordinary green plants is restored and maintained.
The nature and significance of these changes and of the biological activities of which they are the consequence have been the subject of many researches, and are described and discussed in many publications dealing with the soil. Some of these, as for example Charles Darwin's classical account of the activities of earthworms, are familiar to everyone with any interest at all in natural history. As Darwin long ago pointed out, "the activities of earthworms not only operate as a form of natural ploughing, aerating and intimately mixing the soil constituents, but also bring to the surface quantities of humus in forms acceptable to other members of the soil population thus greatly increasing the amounts of nitrogenous and mineral nutrients available for ordinary plants'.
According to recent estimation by American workers, worm castings left on the surface contain five times as much phosphate, and eleven times as much potash in a condition suitable for absorption by roots as does the upper six inches of the soil on which they appear. Realization of the fact that soil fertility can be markedly raised by increasing the worm population has already resulted in the establishment in America of 'worm farms' for the provision of increased supplies of worms appropriate for this purpose.
The researches that revealed the complicated and interlocking life processes of certain soil bacteria, by means of whose activities a supply of nitrates, suitable and sufficient for the needs of ordinary green plants, is maintained in the soil, and those that tell how the vast store of gaseous nitrogen in the atmosphere, useless to higher plants, is drawn upon by certain groups of micro-organisms to build up nitrogen-containing compounds and so brought into circulation, have already been mentioned. Scientific study of the soil has made possible the telling of these fascinating stories, affording instructive glimpses of the hidden life that sustains and renews soil fertility.
Rather less is known in detail of the vital activities of the fungi that live in soil. Various methods have been devised for estimating their numbers and identifying them, all of which have serious drawbacks in respect to the accuracy of the results obtained, and none of which can be trusted to provide reliable information about the group of fungi with which we are here specially concerned. Although there are obvious difficulties in estimating the number of fungi present in soil, it may be taken for granted that very large numbers are present in the upper layers of all fertile soils. Some of the older estimates are undoubtedly much too high; as, for example, that of 1,500,000 fungi in every gramme of soil. Consideration of the ways in which organic material is distributed in a natural soil, dead roots, fallen leaves, and the like, suggests that fungi using as food the materials of which these are composed will tend to congregate where this food is most abundant and it is therefore unlikely that they will be uniformly distributed.
As to the methods available for identifying the fungi present, one of these involves direct microscopic examination of small samples of soil with the use of special stains to facilitate the recognition of hyphae and spores. This can give at best only a very general idea of the amount of fungal growth present in the area whence the sample was taken and little or no information about the kinds of fungi producing it.
The method most commonly used is the same as that adopted for estimating the numbers and kinds of bacteria present in soil and involves the use of ordinary bacteriological procedure. Shallow covered dishes (Petri dishes) each containing a thin layer of nutrient jelly specially suited for fungus growth are sterilized by steam heat and allowed to cool. Small quantities of fresh soil shaken up in known amounts of sterilized water are diluted and then spread evenly over the surface of the jelly, taking the usual precautions to avoid contamination by organisms present in the air. Any fragments of hyphae thus distributed may, if the nutrient provides congenial food material, each give rise to a separate growth or colony which can then be isolated, identified, and studied independently.
In this way large numbers of fungi living in the soil have been observed and placed on record. Some of these are only casual visitors from the air; some are parasites that attack roots and are the cause of troublesome plant diseases; others are saprophytic forms like the familiar moulds.
The importance of the matter from the present point of view lies rather in the negative than in the positive results obtained by these methods, because, curiously enough, the fungi belonging to the group in which we are now interested, although evidently extremely abundant in certain soils, e.g. those of woodlands, and relatively common in almost all soils, are seldom or never isolated in the manner just described, a sufficient comment upon the incomplete character of the evidence provided by these methods, and also incidentally, and more significantly from the present point of view, a reminder that these soil fungi are extremely specialized in respect to their food habits. It is obvious, therefore, that methods for the identification of soil organisms that depend upon the 'sowing' of samples of soil upon specially prepared nutrient surfaces can provide information only about such members of the soil microflora as can use the food substances provided. The absence of particular kinds of fungi or other micro-organisms from such laboratory cultures does not necessarily mean that these were not present in the sample under examination, but only that the food materials presented to them were not acceptable.
What then is really known about the distribution and food habits of the fungi that produce the common toadstools of woodland soils and their relatives on pastures? All belong to the Higher Fungi, a majority to the great group known as Basidiomycetes; a few, for example the truffle fungi and their allies, to the other great subdivision of the Higher Fungi, the Ascomycetes. To the former group belong the Mushroom and all the common 'toadstool fungi' of woodlands. A few of the latter are parasites, attacking and often killing trees; these can hardly be regarded as true soil fungi since they spend the whole or the greater part of their lives within the bodies of their hosts, from which, eventually, the spore-bearing parts emerge (Pl. 1).
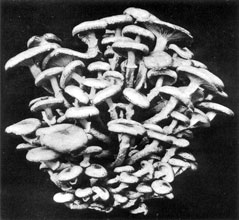
Plate 7. The Honey Agaric, Armillaria mellea. A large cluster of sporophores. About one-third natural size.
|
One of them, the Honey Agaric (Armillaria mellea), is exceptional and shows peculiarities in its mode of life that challenge attention.
A deadly and widely distributed parasite of trees and sometimes of other plants, the Honey Agaric forms its sporophores only after the tissues in which it grows have been killed. The tawny, white-gilled toadstools are common on dead stumps in woodlands and hedgerows, thus betraying the presence of the vegetative mycelium of the parent fungus, at this stage growing as a saprophyte in the dead tissues of the tree and forming root-like strands, that extend into the tissues of the stump and far afield into the soil. These strands or rhizomorphs, looking like dark brown thongs or bootlaces, can grow in soil as well as in living or dead plant tissues, thus serving as a very efficient means of extending the attacks of the parasite to new hosts, spreading from tree to tree through the soil with deadly efficiency. On occasion, the rhizomorphs have been observed to extend into cultivated ground adjoining woodlands, invading potato fields with disastrous consequences to the growing crop. In its mode of life the Honey Agaric is thus not a typical soil fungus. It can be and often is an insidious and dangerous parasite attacking and killing trees and on occasion other kinds of plants; it may grow also as a saprophyte on the dead stumps of its victims, and by means of the rhizomorphs travel long distances through the soil, living upon the organic residues in the humus and thus behaving temporarily like a true soil fungus (Pl. 6, 7, 8, 9).
 Plate 8. The Honey Agaric, Armillaria mellea. Part of the trunk of a tree of Scots pine killed by this fungus, after removal of the bark. The cord-like rhizomorphs travel from the soil up the trunk, growing through the living tissues between the bark and the wood, eventually destroying them and killing the tree. Plate 8. The Honey Agaric, Armillaria mellea. Part of the trunk of a tree of Scots pine killed by this fungus, after removal of the bark. The cord-like rhizomorphs travel from the soil up the trunk, growing through the living tissues between the bark and the wood, eventually destroying them and killing the tree.
|
This remarkable plant has a very wide geographical distribution. In Japan and elsewhere, it is known to form a surprising kind of association with certain orchids of peculiar habit, an association in which the orchid seems to have secured the best of the bargain in respect to food supplies. The first of these associations to be described in detail was that with a curious species of orchid called Gastrodia elata, native in Japan.
The Gastrodia plant when fully developed consists merely of an underground tuber like a large elongated potato or artichoke tapering to one end and covered with brownish cork. The tuber is rootless and without green leaves or shoots; like those of other tubers, the tissues contain reserve food materials, at the expense of which it produces periodically an immense flowering shoot, a yard or more in length, bearing small brownish scale leaves and brownish flowers of the characteristic orchid type (Pl. 9).
It was observed by botanists that the production of flowering shoots by this orchid was very erratic, representing only a small proportion of the mature tubers present in the soil of any given locality. After a time, a Japanese botanist named Kusano made the interesting discovery that the tubers were frequently attacked by rhizomorphs of Armillaria mellea continuous with those infesting cultivated soils or woodlands nearby, and that these rhizomorphs penetrated the tubers by means of sucker-like outgrowths. There followed further researches and experiments leading to the surprising discovery that only tubers parasitized in this way produced flowering shoots, whence it appeared that the mature Gastrodia tuber, cut off from direct contact with the outside world by its corky covering and the absence of roots, re-establishes such contact by suffering parasitic invasion of its tissues. Only thus apparently can it utilize its store of reserve materials to produce flowers and fruit and so complete the life cycle. The details of this curious liaison in the plant world are still obscure. That the fungus gains entry by virtue of its equipment as a parasite seems certain; to what extent it robs the invaded tuber of stored reserve food materials and what it brings in return that enables the latter to utilize those reserves in producing a massive inflorescence giving rise eventually to fruit and seed is still uncertain. It is quite clear, however, that active parasitism of the orchid tuber by the rhizomorphs, if it occurs, is of a very different kind from the ruthless assaults made by the Honey Agaric fungus on potato tubers or living trees. This is in itself a very interesting fact showing how difficult it is to make absolute statements about the food habits of fungi. The relationship of the mycelium with the tissues of the tuber is particularly difficult to explain satisfactorily. There is plenty of suitable food material stored within the tissues and all the mechanism for lethal attack is apparently present in the rhizomorphs. Yet the attack halts at the initial stages, the most obvious result being stimulation of the victim to form new organs and so utilize its own reserves of food!
The Honey Agaric is known to form similar partnerships with another species of Gastrodia and with several East Indian species of orchid having the same curious habit: absence of green leaves or chlorophyll, underground tubers, and the production at intervals of gigantic inflorescences of brownish flowers.
It is known also that another soil fungus, a species of Marasmius, belonging to the same family group as the Honey Agaric, forms a similar kind of relationship with another tropical species of orchid.
All these strange associations are of crucial interest to our present inquiry; first, because they indicate the very wide geographical distribution of a soil fungus such as the Honey Agaric; secondly, and more important, because they indicate what may be called the flexibility of this fungus in respect to its food habits; it can be and usually is an out-and-out parasite on trees or other plants, it can live as a saprophyte in situ upon the dead remains of its victims, or on these and other organic remains in the soil as they pass into the condition of humus. Thirdly, because the orchids involved in these queer relationships are all plants without green leaves or chlorophyll and therefore committed to a mode of life similar to that of their fungus associate. Unable, like green plants, to utilize the inexhaustible supply of carbon present in the air as carbon-dioxide gas, they must seek their essential supplies of this element in organic form as do their fungus competitors. They are indeed, from this point of view, not worth parasitizing!
In what manner the young Gastrodia tuber obtains from the soil the carbon necessary to allow it to reach maturity and convert its tissues into a storehouse of reserve food materials is quite unknown. The impermeable covering of cork acquired subsequently by the mature tuber certainly brings any such absorption to an end, severing completely any direct relations with the soil around it.
As will be learned presently, many -- possibly all -- of the woodland Basidiomycetes also form highly specialized associations with the roots of trees and shrubs, although in other respects they appear to be true soil fungi, growing saprophytically upon the plant and animal residues in the soil, and thus contributing to the formation of the soil humus. In woodland soils it is not difficult to observe whitish or coloured strands of mycelium of these fungi among rotting leaves and in fragments of wood and to infer the part they are playing in the conversion of these into humus.
In order to understand what is happening, in the soil, it is instructive to watch the changes that take place in a compost heap composed of garden refuse, straw, or similar materials as they break down and the heap shrinks in size.
The changes that go on during the conversion of such a heap into humus are very complicated and are brought about by the life activities of many different organisms with varying food requirements. As in soil, various fungi and bacteria play an important part, and it can be shown that the former are specially active in well-aerated heaps in breaking down the more resistant structures that formed the rigid skeletal parts of the plant remains present. It is often possible to observe a dense network of greyish-white mycelium spreading throughout the substance of the heap while sporophores of different fungi, some of them Basidiomycetes of the group we are considering, appear upon the surface.
If the changes taking place are investigated more closely, it becomes evident that definite phases of the breaking-down process are marked by special types of activity such as that just noted in respect to fungus growth. These phases are distinct and do not recur. They serve as an indication that the conversion of such a compost heap into humus is a very complicated affair brought about in stages by the growth activities of different organisms with different food requirements, those of any one series closely linked by invisible ties with those of others that precede and follow them. In course of this conversion the temperature of the heap rises rapidly, often to high levels, due to rapid increase in the numbers of micro-organisms and the chemical changes associated with their intense respiration. Something very similar must be happening more slowly in the soil during the conversion of the raw materials derived from plant and animal remains into humus.
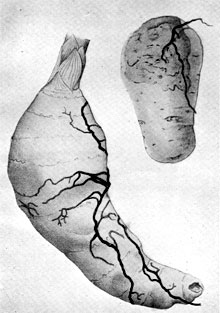 Plate 9. The Honey Agaric, Armillaria mellea. (a) A full-grown tuber of the orchid Gastrodia elata attacked by rhizomorphs of the fungus. The fungus establishes a balanced relationship with the tuber which then produces flowers and fruits. (b) A tuber of potato attacked by rhizomorphs of the same fungus. In this case the relation is purely parasitic and the tuber is eventually destroyed by the fungus. About natural size. Plate 9. The Honey Agaric, Armillaria mellea. (a) A full-grown tuber of the orchid Gastrodia elata attacked by rhizomorphs of the fungus. The fungus establishes a balanced relationship with the tuber which then produces flowers and fruits. (b) A tuber of potato attacked by rhizomorphs of the same fungus. In this case the relation is purely parasitic and the tuber is eventually destroyed by the fungus. About natural size.
|
After passing through the bodies of earthworms or insects when these are present, such residues are a source of food for multitudes of microscopic members of the soil population, among which are many fungi, including those that produce the common toadstools of our woodlands and pastures.
A number of these fungi have been isolated by 'planting out' fragments of young sporophores on suitable nutrients with due precautions as to aseptic procedure. Something has been learned of their food requirements and preferences by transferring mycelium from the colonies of hyphae that grow out from such cultures to various artificial media; a few of them have even been induced to form sporophores under conditions of artificial culture in the laboratory.
In general, however, not a great deal is known in detail about the exact food requirements of these soil fungi when growing under natural conditions, beyond the fact that they are certainly highly specialized in their habits. It is known, for example, that the common mushroom, mycelium of which grows naturally in the soil of pasture fields, can use as a source of food lignin, the rather resistant substance that gives its character to wood, and the same is almost certainly true of many relatives of the mushroom living in woodland soils. It is likely that many of the latter can use as raw materials for food, remains of plants such as twigs and leaves as well as the products of the earlier stages of their disintegration, breaking them down into chemically simpler bodies, although about the details of this there is some disagreement among those who make a special study of soil activities. Even in the case of the common mushroom, there are still considerable gaps in knowledge. Much of the routine practice of mushroom-growing is still empirical, and it is only quite recently that it has been found possible to make composts from straw and other materials to replace satisfactorily the traditional methods of cultivation on stable manure covered with soil. It is also uncertain to what extent and in what manner woodland fungi can make use of humus products in a more advanced stage of decomposition, although it is generally believed that they do use these products, their nutritional activities probably representing an important advance in the condition of these materials towards final disintegration and decay.
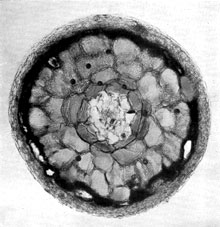 Plate 10. The main features of an ectotrophic mycorrhiza as seen in a rather thick transverse section of a young fungus-root of pine. The photograph shows: the central region of the root not affected by the fungus; the cortex of rather large cells with their intercellular net of mycelium, seen in surface view and also in section between individual cells; cells of the outer layer of the cortex filled with dark tannins; the enveloping 'mantle' of interwoven fungus hyphae continuous with those in the net. Plate 10. The main features of an ectotrophic mycorrhiza as seen in a rather thick transverse section of a young fungus-root of pine. The photograph shows: the central region of the root not affected by the fungus; the cortex of rather large cells with their intercellular net of mycelium, seen in surface view and also in section between individual cells; cells of the outer layer of the cortex filled with dark tannins; the enveloping 'mantle' of interwoven fungus hyphae continuous with those in the net.
|
Certain facts that have been discovered are of great interest as, for example, the observation that a fungus (Coniophora cerebella) common in woodland soils under pines, spruces, and other conifers, in contributing to the breakdown and final decomposition of the conifer needles and other residues that accumulate under the trees, attacks the materials of which these are composed in such a manner as to produce substances of a definite character. These are of such a chemical nature that they cannot be attacked by -- in other words, cannot serve as sources of food material for -- another fungus (Clitocybe nebularis) equally common in deciduous and mixed woodlands but not found in coniferous woods. It is facts of this kind that point the way towards a better understanding of the ways in which the different kinds of woodland soils that are formed under different kinds of trees originate, and indicate how the differences may be directly related with the vital activities of the woodland fungi and other micro-organisms living in them. They indicate why, for example, the organic materials in soils under coniferous trees remain relatively slightly decomposed so giving rise to what are commonly called raw humus soils, while those in deciduous woodlands break down much more thoroughly to form what is called a woodland, mull soil. These soil differences are in turn interrelated with the growth of the trees themselves and made manifest in a very striking way by the toadstools representing the different kinds of woodland fungi that appear beneath them.
In all this, the important thing is to recognize the interlocking and interdependence of the various activities going on in the soil. One man's meat is another man's poison! Organic remains rejected or eliminated by one kind of organism become the food supplies of others. Hence, changes in the soil population or in the kinds of organic residues entering the soil, may have far-reaching consequences upon the character of the humus and may ultimately react upon the fungus flora and even threaten the growth or existence of plants composing the surface vegetation. As an example of this may be cited the disappearance of many common woodland toadstools from areas where the trees have been felled, and their replacement by others produced by quite different fungi.
Trying to focus clearly in our minds all these facts relating to soil fungi and their activities, we realize that they must form an important aspect of the ecology of plants.
Plant ecology is the branch of botanical science concerned with the study of plants in their natural surroundings. The plant ecologist studies the behaviour of plants in relation with all the factors that make up their surroundings in nature. It is easy to distinguish the more important of these that control the physical environment of plants: climatic factors, such as rainfall and range of temperature, winds and weather, with such minor factors as slope of the ground, exposure to or shelter from wind; physical soil factors, related to aeration, water-supply, and the sources of mineral nutrients.
It is rather more difficult to recognize and interpret such soil factors as are determined by the behaviour of other living organisms -- biological factors as they are called. Among the causes of such factors are competition for space and nutrients among members of the ground vegetation, often severe when roots are competing in the same layer of soil, attacks by grazing animals, or interference by man.
It is more difficult still to distinguish clearly those potent factors brought into play through the manifold activities of members of the soil population. If we confine ourselves to the more obvious of the latter depending upon fungal activities, we have learned that mycelium of different kinds of fungi species is living and growing in the soil in close proximity to roots of the plants that form the surface vegetation. This mycelium competes with the roots for water and mineral salts, including such inorganic sources of nitrogen as nitrates and salts of ammonium. It carries on also a multitude of activities in the organic materials present, bringing about chemical changes that condition the character of the humus, so reacting indirectly on the growth of other micro-organisms and on that of the higher plants. The mycelium of some soil fungi may be a source of potential danger to roots, capable of forcing an entry and then parasitizing and even destroying the tissues; the mycelium of others or possibly even of the same species under special conditions may enter the roots or other organs and establish with them regular and constant relationships. Plant ecologists as a whole have tended to overlook the importance of biological factors belonging to this last group, inclining to treat them rather as accidental and casual happenings than as regularly recurring events in the ecological relationships of plants that compose the surface vegetation with members of the soil population -- relationships often of great importance in conditioning the healthy growth of trees and other plants. Some of these associations, as will be learned presently, are significant signposts pointing the way towards an answer to the last question posed at the beginning of our inquiry.
The attempt to find answers to our first two questions, 'What are toadstools? What is their manner of life and how, if at all, does it differ from that of trees and other plants among which they grow?', has led us rather far afield. Knowledge acquired by the way on these excursions has placed us in a better position to seek an answer to the third question, 'Is there any explanation for their distribution in nature; in particular, for the constant association of some kinds with certain species of trees or other plants?'
Next chapter
Back to the Small Farms Library Index

Community development | Rural development
City farms | Organic gardening | Composting | Small farms | Biofuel | Solar box cookers
Trees, soil and water | Seeds of the world | Appropriate technology | Project vehicles
Home | What people are saying about us | About Handmade Projects
Projects | Internet | Schools projects | Sitemap | Site Search | Donations | Contact us
|