Chapter Four
Problems and Partnerships
'He that has partners, has masters.'
-- Attributed to Pope Sixtus V: 1585-92.
There is an Oriental proverb that runs somewhat as follows: Show me a man with very long legs and another with very long arms; together they can pull down the moon. Much of the essence of partnership is contained in this saying: a common goal with diverse and complementary means for reaching it. Yet true partnership implies rather more inasmuch as it involves a period of relative stability or balance, however short, in the mutual relations of the partners.
The contents of the preceding chapters have led us to realize that a partnership of some kind exists between trees and their root associates, the toadstool-forming fungi of woodland soils. It is certain that in nature, trees and fungi pursue a common goal -- the wherewithal to live, grow, and reproduce their kind; certain also that their food requirements are satisfied in different ways. Owing to fundamental differences in the mechanism of nutrition in green and non-green plants, these ways are often complementary to one another when regarded as parts of a general nutritional pattern into which all living things can be fitted.
A word concerning plant partnerships in general. These are common enough; sometimes easy to recognize as such, sometimes not so easy, yielding up their secrets only as a result of elaborate and painstaking researches.
There are partnerships between green and non-green plants; good examples of such are lichens, and trees or other green plants with fungus-roots. A lichen is a composite plant consisting of colourless fungal hyphae more or less closely interwoven to form a compact structure or thallus, imbedded in which are colonies of green cells belonging to members of the simplest green plants or Algae. In respect to sources of food, the two partners are here obviously in similar case to the tree or other green plant possessing fungus-roots. The inherent possibilities of this kind of partnership in respect to nutrition are obvious, if we look no further than the fact that atmospheric carbon-dioxide can be utilized by the green plant but not by the fungus, whereas many organic compounds not directly available to a green plant as food are readily accessible to a fungus. On the other hand, there are partnerships between non-green plants in which the significance of the relation from the nutritional standpoint is far from obvious. Representative of this kind of association are the so-called saprophytic flowering plants, the Bird's Nest Orchid (Neottia nidus-avis), a native British orchid fairly common in certain kinds of woodlands, and the Indian Pipe (Monotropa Hypopitys) usually found growing in beech woods. Both these plants produce clusters of brownish-yellow stems bearing small scale leaves, each stem terminating in a raceme of waxy yellowish flowers. Both are without chlorophyll and form fungus-roots which, in common with the underground stems or rhizomes, are heavily infected with fungus mycelium.
These plants and others like them are commonly referred to by botanists as saprophytes, a name involving the assumption that they can use as sources of food the many and diverse materials locked up in the form of decomposing organic remains in the soil humus. Whether any of them can do this directly is quite uncertain; it is usually presumed that their fungus partners play an important role in such activities, although little is known about the details of nutrition or the mechanism of exchange of nutrients, if this takes place. The strange association described in an earlier chapter between the rootless tuber of an orchid, Gastrodia, and rhizomorphs of the parasitic fungus Armillaria mellea, is perhaps, of all these partnerships between non-green plants, the most difficult to explain.
Another and more unusual kind of partnership is that between a simple animal and a green plant. An example of this is the curious green marine worm, Convoluta roscoffensis, the chief partner in which starts life in the way normal to its kind, as the embryo of a colourless worm, becoming green later as a result of invasion of the tissues by a lowly green plant living in sea-water, closely allied to the green algal partner in many lichens. Colonies of this curious little green animal occur on the beaches of Brittany. At low water and during daylight, the worms come to the surface forming green patches on the sands; when the returning tide reaches them, they submerge again below the surface. By these periodic movements, the green cells within the body are exposed at intervals to sunlight and so enabled to exercise their function of photosynthesis, forming starch from the carbon-dioxide of the atmosphere and so bringing into the body of the worm and placing ultimately at its disposal a supply of food material derived from a source of carbon quite inaccessible to the animal partner alone. The algal partner has at its disposal certain nitrogenous substances formed as waste products in the course of the ordinary life processes of the worm.
All these partnerships with many others are often described as cases of Symbiosis (Gk. sum, with; bioun, life) a word meaning simply, as is evident from its derivation, living with or together. The origin and history of this word is as follows: In the year 1879 a distinguished botanist, Anton de Bary, stimulated by his researches on lichens and greatly impressed by the prevalence in nature of close and often apparently intimate associations between different kinds of organisms, coined a new word, symbiosis, to describe such partnerships in the widest sense.
As used by de Bary, symbiosis meant just what the word implies -- living together. It was intended to cover all cases involving mutualistic relations between two or more different organisms, whether intimate or remote, regardless of the underlying nutritional significance. There would be included, therefore, not only the cases already cited: lichens, mycorrhizas of all kinds, the green worm Convoluta, and the like, in all of which the union of the partners is so complete that to casual inspection there is betrayed no evidence of dual origin, each apparent individual simulating a single plant, plant-member, or animal, as the case may be, but likewise numberless others: parasites with their hosts, flowers with their special pollinating insects, and so forth. Indeed, de Bary drew special attention to the status of parasite and host which he described as 'the best known and most exquisite expression of symbiosis'. ('Die bekannteste und exquisiteste Erscheinung der Symbiose ist der vollständige Parasitismus.')
Modern usage has tended to reject de Bary's definition of symbiosis, and as now used the term almost invariably implies, not merely living together on the part of different organisms, but the existence of reciprocal relations of a kind beneficial to one or both partners. Whether this change in meaning is desirable or not may remain a matter of opinion. It is, however, of some importance that in any given case the reader should appreciate the exact sense in which the term is used, for symbiosis is a rather hard-worked word, apt to be loosely applied in popular accounts of nature studies.
This historical digression will assist the reader to decide for himself, on the evidence provided, whether or not the partnership between trees and toadstools exemplified in the habit of forming fungus-roots or mycorrhizas can properly be described as symbiosis, using the word in the somewhat perverted modern sense to imply mutualistic relations of a beneficial kind.
That mycorrhizal associations in general are all cases of symbiosis in the strict de Bary sense is, of course, evident enough. It is easy also for even the unscientific reader to realize that the expressions 'reciprocal relations' or 'mutualistic' may cover very different kinds of partnerships, ranging from that in which one partner gets most of the benefit to others in which there is a give-and-take relation of a most delicately adjusted kind; in which maintenance of a condition of equilibrium depends upon alternation of successive phases, during which now one now the other partner benefits.
What we require to know is whether one or other of these kinds of partnership is represented in the trees that form fungus-roots with the toadstool-forming fungi of woodlands.
It is needless to labour the point that every possible opinion has been held by botanists respecting the physiological relations of the partners in mycorrhizal plants; needless also to recapitulate the arguments, often fallacious, on, which these opinions were based.
Controversy initiated nearly sixty years ago by certain of Frank's contemporaries has found an echo recently in an opinion expressed by a modern botanist: 'The presence of the fungi in a mycorrhizal association is to be regarded as an example of controlled parasitic attack and has no mutualistic significance.' Discussing the uniformity of structure observed in mycorrhizas in respect to the details of fungus infection, Burges further remarks that this 'is probably no more than the result of the uniform conditions in the soil leading to a uniform degree of disease'. (The italics are the writer's.)
Let us examine our special case of trees for evidence bearing on this, taking the Scots pine (Pinus silvestris) as a typical example, one also in which the identity of many of the fungus associates is known. It is beyond dispute that the healthier and more vigorous a pine tree under natural conditions, the more profusely is the root system furnished with mycorrhizas. This is readily confirmed by examination of seedlings appearing naturally beneath parent trees or growing in nurseries or pot-cultures. Not only is it an established fact that healthy seedlings are always well furnished with short roots all of which become mycorrhizas, but it is evident that the number of the latter increases with the size and vigour of the seedlings. There is, indeed, no surer or safer way of assessing the health and vigour of young pine trees, or judging the suitability or otherwise of soil conditions for their growth, than by estimation of the number of mycorrhizas they possess and critical examination of the mycorrhizal structure. Soil conditions unfavourable to growth are at once reflected in reduction in number or complete disappearance of fungus-roots with the development of abnormal structural features; improved soil conditions by increased mycorrhizal activity. There is, in short, no evidence at all to support the view that fungus-roots of pines and other trees of like habit are a symptom of disease. To hold such a view is paradoxical in the extreme. On the contrary, all recent experimental evidence confirms the opinion expressed by the Swedish botanist, Melin, in 1923, that mycorrhizas are of vital importance for seedlings and trees in raw-humus and related soil types. ('Dass die Mykorrhizen für die Pflänzchen und Baüme auf Roh-humusböden und verwandten Bodentypen eine vitale Bedeutung besitzen.')
Frank's conclusions as to the significance of fungus-roots in tree nutrition were based partly on inferences from comparative observations, partly on experimental results. These earlier experiments do not always satisfy modern standards, thus accounting in part for subsequent disagreement and controversy. The general conclusions reached by Frank and the early observers are, however, now strongly reinforced by experimental results obtained in recent researches, leading irresistibly to the following conclusions:
- The short roots of pine, spruce, and trees of similar habit are distinct organs of limited growth in length and relatively short duration, usually functioning for only one year. Under favourable growth conditions, these short roots are immediately subject to fungus infection, becoming mycorrhizas of the ectotrophic type. Whether infected or not, they retain their characters as short roots, recognizable on anatomical grounds alone.
- There is reason to believe that the production of short roots is influenced favourably by the presence of active mycelium of the mycorrhiza-forming fungi in the soil about them, possibly to a less extent by that of other soil fungi.
- Mycelium of the root-fungi does not normally 'attack' the long roots of the host. Whenever it does so, it is a sure indication of the presence of disturbing soil factors.
- Short roots that for any reason do not become mycorrhizas are often subject to attack by other soil fungi, with deleterious effects on the health and growth of the trees. They thus become pseudo-mycorrhizas, usually readily distinguishable by their external appearance and always entirely different from true mycorrhizas in structure.
- Failure to form mycorrhizas may be due to unfavourable soil factors that react adversely on the condition of the roots or the mycelium, so inhibiting normal association, or to absence of appropriate mycelium.
- There is conclusive experimental evidence that normal healthy and vigorous growth of the host trees follows upon and is directly related with the free development of mycorrhizas.

Plate 15. Fungus-roots of Norway Spruce (Picea abies). The branching resembles that of fungus-roots of beech (see Pl. 11), quite unlike the repeated forking into two equal branches of the mycorrhizas of pine. (See Pl. 12.)
|
The photograph shows part of the rooting area of seedlings of this pine grown in a glass-fronted box. The soil and seed used were sterilized and the mycelium of the fungus introduced by inoculation of the soil from a pure culture. Slightly magnified.
If these conclusions are accepted, it is evident that they must be of great importance in forestry, especially in nurseries and afforestation areas on soils from which the appropriate woodland fungi are absent; as, for example, on the prairie soils of the middle-western states of the United States; also in the establishment of trees not native in the country or locality concerned.
As not infrequently happens, recognition of this in practice anticipated scientific proof of the exact causes at work, as numbers of more or less empirical experiments carried out by practical foresters in the course of forestry operations in different parts of the world bear witness, For example, it is frequently found difficult to raise satisfactory seedlings of pine, spruce, and the other trees concerned in new nurseries situated on soils of non-woodland type and distant from woodlands.
The seedlings from such nurseries have poor root systems deficient in short roots and mycorrhizas, with stunted shoots bearing yellowish or purplish-brown needles. These undesirable effects on growth are removed by importing to the new nursery seedlings of the same species or one closely allied from a woodland nursery, furnished with normal mycorrhizas, and allowing them to grow in the same beds for a season preceding seed-sowing. This procedure has become part of the routine of ordinary nursery practice and is now correctly interpreted as indicating the necessity for introducing suitable mycorrhizal fungi.
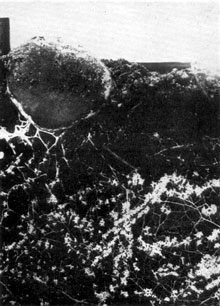
Plate 16. A sporophore of Rhizopogon luteolus developing near mycorrhizas of Pinus taeda, formed by association with this fungus in an experimental culture.
|
By carrying out more exact experiments in pots or boxes with soil treated by heat or antiseptics in such a way as to free it from fungal growth of any kind, and using seeds washed in antiseptics to remove surface contamination, it is easy to raise seedlings of pine and similar trees free from mycorrhizal or other fungus infection and then to compare their growth with that of others grown in precisely the same way, but supplied with fungus-roots following the introduction of mycelium of suitable fungi to the seed bed.
The presence of appropriate mycelium can be secured by more exact methods than that just described, by introducing very small quantities of humus known to contain the desired mycelium, or better still, by inserting in the soil small fragments of a sporophore or mycelium from a pure culture of the fungus desired.
It is worthy of note that in all experiments of this kind in the course of which inoculating material is introduced into pots or seed beds as separate fragments, the formation of fungus-roots with the associated improvement in growth of the seedlings begins in plants at or near the points of inoculation, whence it spreads outwards throughout the bed or pot.
Similar experiments of this kind, using various tree species, have been carried out in different parts of the world with identical results. The photographs reproduced relate to experiments with Corsican pine in Great Britain (Pl. 17), to a Himalayan species of pine, Pinus longifolia, in a nursery bed in Rhodesia [Zambia] (Pl. 18), and to a North American species, Pinus taeda, in Queensland, Australia (Pl. 16).
The experiment illustrated in the very beautiful photograph reproduced in Plate 16 was carried out in a glass-fronted box using sterilized soil and seed. The fungus introduced had been observed in field plantations of the same pine in Queensland and was isolated from a young sporophore and grown in pure culture in the laboratory. Eight months after sowing the pine seeds, fragments of mycelial growth from such a culture were inserted in the soil of the boxes; profuse development of mycorrhizas followed, starting in each case at the point of insertion. About three months later sporophores of the fungus were formed in the neighbourhood of the mycorrhizas either below soil level, the normal position, or at the surface, as shown in the photograph. The fungus in question, Rhizopogon luteolus, is a Gasteromycete, closely related to the common puff-ball fungi, and occurs not infrequently in Great Britain. Its appearance in Queensland and elsewhere in Australia is an example of the very wide geographical distribution of these mycorrhiza-forming fungi of trees.
In view of the intricacy of the subject and limitations of space, it is possible to deal only very briefly with the many interesting facts that have emerged from recent experimental work of this kind. If the short review just offered has persuaded the reader that the association between trees and toadstools can properly be described as symbiotic in the sense that it involves mutual relations of a kind beneficial to the healthy growth of either or both partners, he is entitled to ask whether there is any positive evidence about the existence of a mechanism whereby exchange of nutrients can take place.
The older views about this are associated chiefly with the names of two German botanists, Frank and Stahl. As already mentioned, Frank believed that the mycelium of the root-fungi brought to the roots not only water and salts, but also soluble organic food materials, in particular those containing nitrogen derived from the soil humus. Later, Stahl emphasized the great importance of mycorrhizas in increasing the efficiency of absorption by the roots, but held that their chief importance lay not so much in respect to acquisition of nitrogenous food as in their capacity for absorbing mineral salts and in making use of non-nitrogenous organic compounds. In neither case was the experimental evidence conclusive in respect to these claims. Modern work supports Stahl's interpretation by providing proof of the greatly increased absorbing surface offered by a well-developed, mycorrhizal system, also some direct evidence that the fungus mantle acts as a protection against damage to the root from severe drought. It has been observed also that the concentration of mineral salts in the soil water increases in the zone of soil immediately surrounding the mycorrhizas, and there is some evidence of a rather theoretical kind that the presence of mycelium within the root tissues facilitates the absorption of soluble nutrients from the soil.
Direct experimental evidence that fungus-roots can absorb soluble organic substances derived from the soil humus is scanty and not very satisfactory in character, and there is still no exact knowledge about actual exchange of food materials within the roots.
On the whole, the results of modern researches tend to confirm the older views in so far as they stress the importance of fungus-roots in tree nutrition. Botanists have been inclined to regard these older views -- as put forward by Frank and Stahl, for example -- rather as a battle-ground for controversy than as useful contributions to the solution of an extremely complex problem.
A more advanced stage in the study of mycorrhizal relations in trees has now been reached. It appears to the writer that there is no need to regard the theories of nutrition championed by Frank and Stahl respectively as mutually exclusive. On the contrary, each may find a place within the wide range of conditions known to exist.
There is now available reliable information about another aspect of nutrition unsuspected by the earlier workers. In addition to the rather elusive substances known as vitamins so important in animal nutrition, there are produced by plants definite chemical substances included under the general name of growth-promoting substances or auxins. For example, many fungi produce a substance, known to botanists as heteroauxin and to chemists as indole-acetic acid, that stimulates the production of roots, although it has a retarding effect on their growth in length. This is a fact of great significance in view of the activities of fungus mycelium in the soil, and the profuse formation of short roots in seedling trees that precedes the conversion of these to mycorrhizas under favourable conditions. On the other hand, evidence has long been available that substances stimulating to the growth of fungus mycelium are excreted by young tree roots; recent work suggests that the exudates from the roots may act in this way because they contain definite growth-promoting substances required by the fungus. Indeed, a Swedish investigator has recently expressed the opinion that interchange of growth-promoting substances rather than of ordinary nutrients is the most important activity in the mycorrhizal relations of trees.
In what other ways, if any, the fungus partner may benefit from the association is at present uncertain. The undoubted fact that many of these fungi do not form sporophores and therefore do not complete their life-cycle except when functioning as root-fungi in association with trees, suggests very strongly that beneficial effects on growth are not confined to the trees but are reciprocal in character.

Plate 17. Seedlings of Corsican pine (Pinus nigra), one year old, showing the effects upon the growth and size of the plants that followed addition to the seed bed of a very small quantity of rootlets and soil from the Corsican forest of ïtoneA, where this tree is native. Left: seedlings from bed to which Corsican humus was added. Right: seedlings from adjoining bed of similar soil without this addition.
|
It may be asked whether the problems of nutrition presented by the mycorrhizal habit in trees can be solved by the method of pure culture already described. If tree seedlings can be grown under complete control with and without their fungus associates, should it not be possible by varying the conditions to find out a good deal about the nutritive relations of the two partners?
This brings us to the crux of the whole matter. In nature, fungus-roots are formed and carry on their activities in the soil, a highly complicated and constantly shifting environment, difficult, indeed impossible to reproduce under pure culture conditions. It is not surprising, therefore, that this method of experiment may yield results and lead to interpretations quite at variance with those from experiments conducted under natural conditions.

Plate 18. Two beds of seedlings of a Himalayan species of pine, Pinus longifolia, from the same sowing in Northern Rhodesia [Zambia]. Further bed inoculated with soil containing a mycorrhiza-forming fungus for the species from a vigorous plantation of this pine at a station in Southern Rhodesia [Zimbabwe] 1000 miles distant; nearer bed untreated.
|
The mycorrhizal habit must have originated as a mild attack upon roots by soil fungi living in competition with them, and it appears certain must have had a long evolutionary history. If so, it might be expected that different stages of evolution would be represented among living plants, varying in respect to details of the nutritive relations, to the character of the profit-and-loss account for each partner, and to the degree of stability represented in the partnership. In the special case now under consideration, the constancy of the habit and the remarkable uniformity of structure in the fungus-roots formed by trees in association with the mycelium of certain fungi of woodland soils, suggest that the characteristic features of such soils are in some way directly concerned; suggest, in fact, that the formation of fungus-roots by these trees does not take place as an isolated activity, but is very definitely bound up with other soil factors.
The multiple habit in trees, namely, that of forming associations with a number of different fungi alternatively, reinforces this point of view, suggesting as it does that association with the mycelium of one fungus species rather than another may be determined by the soil environment, not only because this may in itself limit the number of fungal species available, but because it may determine which if any of these species is capable of forming a stable and useful association under the existing soil conditions.
It appears possible that much of the confusion and controversy provoked by study of the mycorrhizal habit in trees and other plants may have been caused by viewing it as an isolated botanical phenomenon, a speciality of certain kinds of plants, rather than as a response on the part of these plants to definite factors in their surroundings.
Viewed in this way, it is evident that in any given soil, the fungi present and capable of forming fungus-roots with trees growing naturally on that soil or introduced by man, must be regarded as soil factors of great importance.
Recent experimental work provides support for this ecological point of view. It is known, for example, that the details of mycorrhizal equipment -- the number of fungus-roots formed and their structural characters -- correspond with remarkable delicacy with natural and artificially induced changes in the soil environment. It has been learned also that fungi harmless to a given species of tree under native conditions may attack the roots and damage the growth of the same species when transferred to another soil; in the first case, there is a balanced condition of equilibrium in respect to the activities of the soil population; in the second, this balance is disturbed.
Considerations of this kind lead to a new point of view, that of the plant ecologist. As already pointed out, plant ecology is the branch of botany concerned with the study of plants in relation with their environment. Ecologists are aware that changes are constantly taking place in the character of the vegetation of any given area. They can register such changes and investigate the factors that operate in bringing them about. Often it is possible to observe definite stages in the course of transition and study in detail any instability in the character of the vegetation brought about by the fluctuating behaviour of one or other of the plant species that compose it.
The natural surroundings that make up the environment can be analysed into a large number of factors, some of which are significant in their effects, others not.
The soil is an important part of the plant's environment and many soil factors -- physical, chemical, and biological -- are directly and effectively concerned. Although much attention has been given to the study of soil factors and their effects upon plant growth and behaviour, there has been a tendency to underrate the importance of biological as compared with chemical and physical causes; in particular, the significance of fluctuations and changes that may take place in the behaviour of the myriad unseen members of the soil population. It seems likely that in any given case, there may be instability in respect to the character of the soil population just as there is instability in that of the surface vegetation. The proper functioning of the fungus-roots and their significance to the host trees may at times be bound up with the maintenance of a condition of ecological stability in the soil. Disturbance of the balanced activities on which this stability depends may affect the character of the mycorrhizal relations with possible reactions upon healthy growth of the trees.
Not a great deal is known in detail about changes in the character of the soil population, or of how the balance of power among its members is maintained or restored. Fragments of knowledge bearing on this are suggestive; as already noted, it has been observed that mycelium of one kind of woodland fungus in the course of its ordinary nutritive activities may bring about chemical changes in the humus constituents of certain kinds of woodland soils that render the products unacceptable as sources of food to the mycelium of another fungus equally common in the soil of other kinds of woodlands. Knowledge of this kind provides clues towards better understanding of the ways in which different kinds of humus and eventually different kinds of woodland soils are produced.
In quite another sphere there has been recently a remarkable opportunity for learning how the antagonisms and incompatibilities that exist commonly among soil micro-organisms living in close proximity express themselves.
All the world now knows of penicillin and the curative effects that result from its action in arresting the activities of bacteria that cause serious disease in the animal body. Penicillin is produced and excreted by Penicillium notatum, one of the mould fungi. When brought into contact with certain bacteria, it has the effect not of killing them outright but of slowing down their vital activities so that they cease to grow and multiply. There is good reason for believing that Penicillium notatum and its near allies are themselves soil fungi. Substances with similar properties to penicillin -- antibiotic substances as they are called -- are now known to be produced by other members of the soil population; for example, by other species of mould fungi, by some of the woodland fungi that form mycorrhizal associations with trees, by certain bacteria, and by other common soil organisms named Actinomyces.
This new and enthralling field of research has attracted many workers. A number of antibiotic substances other than penicillin formed by micro-organisms have already been prepared and tested; none of those yet found possesses the combination of properties, including low toxicity to human tissues, that renders penicillin so valuable a weapon in medical science.
Apart from this aspect of the subject, these new discoveries help us to understand how extremely complex are the soil conditions that control the ecology of soil fungi, and how delicate is the mechanism by which the balance of power is maintained among members of the soil population. Inasmuch as the growth of trees is affected profoundly by the activities of soil fungi, directly by those parasitic forms that cause root diseases, as well as by the symbiotic species that form mycorrhizas, and indirectly by the interplay of other biological factors that impinge upon these fungal activities, it is evident that disturbance of any of the soil conditions may ultimately react upon the degree of well-being of the trees.
It is helpful and not too far-fetched to think of the soil as itself an organism -- a social organism like a human society, for example -- the manifold vital activities of which are carried on by its numerous living inhabitants. Disturbance of any one of these activities may affect others and thus lead to loss of equilibrium and the appearance of symptoms of disorder, with eventually perhaps an increasing degree of biological inertia of one kind or another, a condition just the reverse of the vital activity characteristic of a living and fertile soil.
THE END
Back to the Small Farms Library Index

Community development | Rural development
City farms | Organic gardening | Composting | Small farms | Biofuel | Solar box cookers
Trees, soil and water | Seeds of the world | Appropriate technology | Project vehicles
Home | What people are saying about us | About Handmade Projects
Projects | Internet | Schools projects | Sitemap | Site Search | Donations | Contact us
|